Cell Injury and Cellular Adaptations
By Devesh Chaudhari
Published on 02-Sep-2020Cells are the basic units of tissues, which form organs and systems in the human body.
Traditionally, body cells are pided in to two main types: epithelial and mesenchymal cells. In health, the cells
remain in accord with each other. In 1859, Virchow first published cellular theory of disease, bringing in the
concept that diseases occur due to abnormalities at the level of cells. Since then, study of abnormalities in
structure and function of cells in disease has remained the focus of attention in understanding of diseases. Thus,
most forms of diseases begin with cell injury followed by consequent loss of cellular function. Cell injury is
defined as a variety of stresses a cell encounters as a result of changes in its internal and external environment.
In general, cells of the body have inbuilt mechanism to deal with changes in environment to an extent.
The cellular response to stress may vary and depends upon the following variables:
i) The type of cell and tissue involved.
ii) Extent and type of cell injury.
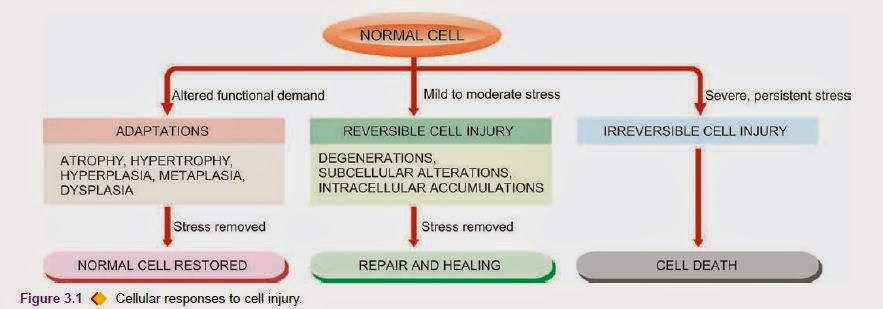
Various forms of cellular responses to cell injury may be as follows
1. When there is increased functional demand, the cell may adapt to the changes which are expressed morphologically
and
then revert back to normal after the stress is removed
2. When the stress is mild to moderate, the injured cell may recover (reversible cell injury), while when the injury
is
persistent cell death may occur (irreversible cell injury).
3. The residual effects of reversible cell injury may persist in the cell as evidence of cell injury at subcellular
level (subcellular changes), or metabolites may accumulate within the cell (intracellular accumulations). In order
to
learn the fundamentals of disease processes at cellular level, it is essential to have an understanding of the
causes
and mechanisms of cell injury and cellular adaptations, which can be best understood in the context of basic
knowledge
of normal structure and functions of cell outlined below.
THE NORMAL CELL Different types of cells of the body possess features which distinguish one type from another. However, most mammalian cells have a basic plan of common structure and function, except the red blood cell which is devoid of nucleus.
CELL STRUCTURE: Under normal conditions, cells are dynamic structures existing in fluid environment. A cell is enclosed by cell membrane that extends internally to enclose nucleus and various subcellular organelles suspended in cytosol Cell Membrane Electron microscopy has shown that cell membrane or plasma membrane has a trilaminar structure having a total thickness of about 7.5 nm and is known as unit membrane. The three layers consist of two electron-dense layers separated by an electron lucent layer. Biochemically, the cell membrane is composed of complex mixture of phospholipids, glycolipids, cholesterol, proteins and carbohydrates. These layers are in a gel-like arrangement and are in a constant state of flux. The outer surface of some types of cells shows a coat of mucopolysaccharide forming a fuzzy layer called glycocalyx. Proteins and glycoproteins of the cell membrane may act as antigens (e.g. blood group antigens), or may form receptors (e.g. for viruses, bacterial products, hormones, immunoglobulins and many enzymes). The cell receptors are probably related to the microtubules and microfilaments of the underlying cytoplasm. The microtubules connect one receptor with the next. The microfilaments are contractile structures so that the receptor may move within the cell membrane. Bundle of microfilaments along with cytoplasm and protein of cell membrane may form projections on the surface of the cell called microvilli. Microvilli are especially numerous on the surface of absorptive and secretory cells (e.g. small intestinal mucosa) increasing their surface area. In brief, the cell membrane performs the following important functions:i) Selective permeability that includes diffusion, membrane pump (sodium pump) and pinocytosis (cell drinking).
ii) Bears membrane antigens (e.g. blood group antigens, transplantation antigen).
iii) Possesses cell receptors for cell-cell recognition and communication.
 Nucleus
Nucleus
The nucleus consists of an outer nuclear membrane enclosing nuclear chromatin and nucleoli.
NUCLEAR MEMBRANE The nuclear membrane is the outer envelop consisting of 2 layers of the unit membrane which are separated by a 40-70 nm wide space. The outer layer of the nuclear membrane is studded with ribosomes and is continuous with endoplasmic reticulum. The two layers of nuclear membrane at places are fused together forming circular nuclear pores which are about 50 nm in diameter. The nuclear membrane is crossed by several factor which regulate the gene expression and repair the DNA damage as soon as it occurs.
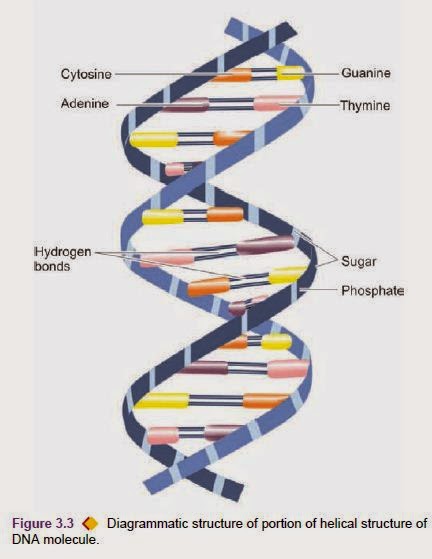
NUCLEAR CHROMATIN The main substance of the nucleus is comprised by the nuclear chromatin which is in the form of shorter pieces of thread-like structures called chromosomes of which there are 23 pairs (46 chromosomes) Schematic diagram of the structure of an epithelial cell. together measuring about a metre in length in a human diploid cell. Of these, there are 22 pairs (44 chromosomes) of autosomes and one pair of sex chromosomes, either XX (female) or XY (male). Each chromosome is composed of two chromatids connected at the centromere to form ‘X’ configuration having variation in location of the centromere. Depending upon the length of chromosomes and centromeric location, 46 chromosomes are categorised into 7 groups from A to G according to Denver classification (adopted at a meeting in Denver, USA). Chromosomes are composed of 3 components, each with distinctive function. These are: deoxyribonucleic acid (DNA) comprising about 20%, ribonucleic acid (RNA) about 10% , and the remaining 70% consists of nuclear proteins that include a number of basic proteins (histones), neutral proteins, and acid proteins. DNA of the cell is largely contained in the nucleus. The only other place in the cell that contains small amount of DNA is mitochondria. Nuclear DNA along with histone nuclear proteins form bead-like structures called nucleosomes which are studded along the coils of DNA. Nuclear DNA carries the genetic information that is passed via RNA into the cytoplasm for manufacture of proteins of similar composition. During cell pision, one half of DNA molecule acts as a template for the manufacture of the other half by the enzyme, DNA polymerase, so that the genetic characteristics are transmitted to the next progeny of cells (replication). The DNA molecule as proposed by Watson and Crick in 1953 consists of two complementary polypeptide chains forming a double helical strand which is wound spirally around an axis composed of pentose sugar-phosphoric acid chains. The molecule is spirally twisted in a ladder-like pattern, the steps of which are composed of 4 nucleotide bases:
two purines (adenine and guanine, i.e. A and G) and two pyrimidines (cytosine and thymine, i.e. C and T); however, A pairs specifically with T while G pairs with C . The sequence of these nucleotide base pairs in the chain, determines the information contained in the DNA molecule or constitutes the genetic code. In April 2003, sequencing of human genome was completed which revealed that 23 pairs of chromosomes in the nucleus of each human cell contains approximately 3 billion base pairs, and each chromosome contains an estimated 30,000 genes in the human genome, which carry the instructions for making proteins. In the interphase nucleus (i.e. between mitosis), part of the chromatin that remains relatively inert metabolically and appears deeply basophilic due to condensation of chromosomes is called heterochromatin, while the part of chromatin that is lightly stained (i.e. vesicular) due to dispersed chromatin is called euchromatin. For example, in lymphocytes there is predominance of heterochromatin while the nucleus of a hepatocyte is mostly euchromatin.
NUCLEOLUS.
The nucleus may contain one or more rounded bodies called nucleoli. Nucleolus is the site of synthesis of ribosomal RNA. Nucleolus is composed of granules and fibrils representing newly synthesised ribosomal RNA. Cytosol and Organelles The cytosol or the cytoplasm is the gel-like ground substance in which the organelles (meaning little organs) of the cells are suspended. These organelles are the site of major enzymatic activities of the cell which are possibly mediated by enzymes in the cytosol. The major organelles are the cytoskeleton, mitochondria, ribosomes, endoplasmic reticulum, Golgi apparatus, lysosomes, and microbodies or peroxisomes.
1. CYTOSKELETON. Microfilaments, intermediate filaments, and microtubules are responsible for maintaining cellular form and movement and are collectively referred to as cytoskeleton.
i) Microfilaments are long filamentous structures having a diameter of 6-8 nm. They are composed of contractile proteins, actin and myosin, and perse materials like parts of microtubules and ribonucleoprotein fibres. Bundles of microfilaments are especially prominent close to the plasma membrane and form terminal web. Extension of these bundles of microfilaments along with part of plasma membrane on the surface of the cell form microvilli which increase the absorptive surface of the cells.
ii) Intermediate filaments are filamentous structures, 10 nm in diameter, and are cytoplasmic constituent of a number of cell types. They are composed of proteins. There are 5 principal types of intermediate filaments:
a) Cytokeratin (found in epithelial cells).
b) Desmin (found in skeletal, smooth and cardiac muscle).
c) Vimentin (found in cells of mesenchymal origin).
d) Glial fibrillary acidic protein (present in astrocytes and
ependymal cells).
e) Neurofilaments (seen in neurons of central and peripheral nervous system).
Their main function is to mechanically integrate the cell organelles within the cytoplasm.
iii) Microtubules are long hollow tubular structures about 25 nm in diameter. They are composed of protein, tubulin. Cilia and flagella which project from the surface of cell are composed of microtubules enclosed by plasma membrane and are active in locomotion of the cells. Basal bodies present at the base of each cilium or flagellum and centriole located at the mitotic spindle of cells are the two other morphologically similar structures composed of microtubules.
2. MITOCHONDRIA.
Mitochondria are oval structures and are more numerous in metabolically active cells. They are enveloped by two layers of membrane—the outer smooth and the inner folded into incomplete septa or sheaf-like ridges called cristae. Chemically and structurally, membranes of mitochondria are similar to cell membrane. The inner membrane, in addition, contains lollipop-shaped globular structures projecting into the matrix present between the layers of membrane. The matrix of the mitochondria contains enzymes required in the Krebs’ cycle by which the products of carbohydrate, fat and protein metabolism are oxidised to produce energy which is stored in the form of ATP in the lollipop-like globular structures. Mitochondria are not static structures but undergo changes in their configuration during energised state by alteration in the matrix and intercristal space; the outer membrane is, however, less elastic. Mitochondria perform the important metabolic function of oxidative phosphorylation, and in the process generate free radicals injurious to membranes. They also have role in apoptosis. Mitochondria contain 37 genes out of which 13 encode for synthesising proteins. In addition, mitochondria also have some DNA and ribosomes.
3. RIBOSOMES.
Ribosomes are spherical particles which contain 80-85% of the cell’s RNA. They may be present in the cytosol as ‘free’ unattached form, or in ‘bound’ form when they are attached to membrane of endoplasmic reticulum. They may lie as ‘monomeric units’ or as ‘polyribosomes’ when many monomeric ribosomes are attached to a linear molecule of messenger RNA. Ribosomes synthesise proteins by translation of messenger RNA into peptide sequences followed by packaging of proteins for the endoplasmic reticulum.
4. ENDOPLASMIC RETICULUM.
Endoplasmic reticulum is composed of vesicles and intercommunicating canals. It is composed of unit membrane which is continuous with both nuclear membrane and the Golgi apparatus, and possibly with the cell membrane. The main function of endoplasmic reticulum is the manufacture of protein. Morphologically, there are 2 forms of endoplasmic reticulum: rough (or granular) and smooth (or agranular).
i) Rough endoplasmic reticulum (RER) is so-called because its outer surface is rough or granular due to attached ribosomes on it. RER is especially well-developed in cells active in protein synthesis e.g. Russell bodies of plasma cells, Nissl granules of nerve cells.
ii) Smooth endoplasmic reticulum (SER) is devoid of ribosomes on its surface. SER and RER are generally continuous with each other. SER contains many enzymes which metabolise drugs, steroids, cholesterol, and carbohydrates and partake in muscle contraction.
5. GOLGI APPARATUS.
The Golgi apparatus or Golgi complex is generally located close to the nucleus. Morphologically, it appears as vesicles, sacs or lamellae composed of unit membrane and is continuous with the endoplasmic reticulum. The Golgi apparatus is particularly welldeveloped in exocrine glandular cells. Its main functions are synthesis of carbohydrates and complex proteins and packaging of proteins synthesised in the RER into vesicles. Some of these vesicles may contain lysosomal enzymes and specific granules such as in neutrophils and in beta cells of the pancreatic islets.
6. LYSOSOMES.
Lysosomes are rounded to oval membrane-bound organelles containing powerful lysosomal digestive (hydrolytic) enzymes. There are 3 forms of lysosomes:
i) Primary lysosomes or storage vacuoles are formed from the various hydrolytic enzymes synthesised by the RER and packaged in the Golgi apparatus.
ii) Secondary lysosomes or autophagic vacuoles are formed by fusion of primary lysosomes with the parts of damaged or worn-out cell components.
iii) Residual bodies are indigestible materials in the lysosomes, e.g. lipofuscin.
7. CENTRIOLE OR CENTROSOME.
Each cell contains a pair of centrioles in the cytoplasm close to nucleus in the area called centrosome. Centrioles are cylindrical structure composed of electron-dense evenly-shaped microtubules. They perform the function of formation of cilia and flagellae and constitute the mitotic spindle of fibrillary protein during mitosis.
INTERCELLULAR COMMUNICATION
All cells in the body constantly exchange information with each other to perform their functions properly. This
process
is accomplished in the cells by direct cell-to-cell contact (intercellular junctions), and by chemical agents, also
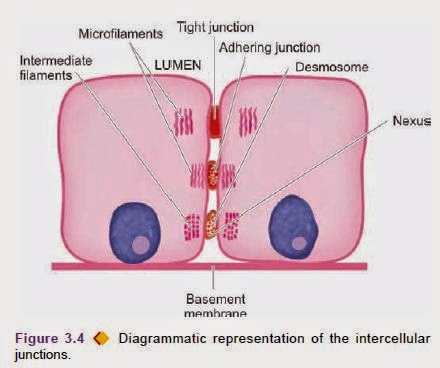
called as molecular agents or factors (molecular interactions between cells) as under. Intercellular Junctions
Plasma
membranes of epithelial and endothelial cells, though closely apposed physically, are separated from each other by
20 nm
wide space. These cells communicate across this
space through intercellular junctions or junctional complexes visible under electron microscope and are of 4 types:
1. Occluding junctions (Zonula occludens). These are tight junctions situated just below the luminal margin of
adjacent
cells. As a result, the regions of occluding zones are impermeable to macromolecules. The examples of occluding
zones
are seen in renal tubular epithelial cells, intestinal epithelium, and vascular endothelium in the brain
constituting
blood-brain barrier.
2. Adhering junctions (Zonula adherens). These are located just below the occluding zones between the adjacent cells
and
are permeable to tracer particles. These zones are in contact with actin microfilaments e.g. in small cell carcinoma
of
the lung.
3. Desmosomes (Macula densa). These are tiny adhesion plates present focally between the adjacent epithelial cells,
especially numerous in the epidermis. Bundles of intermediate filaments (termed tonofilaments in the case of
epidermis)
project from the intercellular desmosomes and radiate into the cytoplasm. Hemidesmosomes are a variant of
desmosomes,
occurring at the basal region of epithelial cells between plasma membrane and the basement membrane.
4. Gap junctions (Nexus). Gap junctions or nexus are the regions on the lateral surfaces of epithelial cells where
the
gap between the adjoining plasma membranes is reduced from 20 nm to about 2 nm in width. Pits or holes are present
in
the regions of gap junctions so that these regions are permeable to small tracer particles. Molecular Interactions
between Cells Besides having intercellular junctions, most cells
communicate at molecular level as follows:
1. Cell adhesion molecules (CAMs)
2. Cytokines
3. Membrane receptors
1. CELL ADHESION MOLECULES (CAMs).
These are chemicals which mediate the interaction between cells (cell-cell interaction) as well as between cells and
extracellular matrix (cell-ECM interaction). The ECM is the ground substance or matrix of connective tissue which
provides environment to the cells and consists of 3 components:
i) fibrillar structural proteins (collagen, elastin);
ii) adhesion proteins (fibronectin, laminin, fibrillin, osteonectin, tenacin); and
iii) molecules of proteoglycans and glycosaminoglycans (heparan sulphate, chondroitin sulphate, dermatan sulphate,
keratan sulphate, hyaluronic acid). CAMs participate in fertilisation, embryogenesis, tissue
repair, haemostasis, cell death by apoptosis and in inflammation. CAMs may be detected on the surface of cells as
well
as free in circulation. There are 5 groups of CAMs:
i) Integrins. They have alpha (or CD11*) and beta (CD18) subunits and have a role in cell-ECM interactions and in
leucocyte-endothelial cell interaction.
ii) Cadherins. These are calcium-dependent adhesion molecules which bind adjacent cells together and prevent
invasion
of ECM by cancer cells. Various types of cadherins include: E-cadherin (epithelial cell), N-cadherin (nerve cell),
M-cadherin (muscle cell), and P-cadherin (placenta).
iii) Selectins. Also called as lectins, these CAMs contain lectins or lectin-like protein molecules which bind to
glycoproteins and glycolipids on the cell surface. Their major role is in movement of leucocytes and platelets and
develop contact with endothelial cells. Selectins are of 3 types:
P-selectin (from platelets, also called CD62), E-selectin (from endothelial cells, also named ECAM), and L-selectin
(from leucocytes, also called LCAM).
iv) Immunoglobulin superfamily. This group consists of a variety of immunoglobulin molecules present on most cells
of
the body. These partake in cell-to-cell contact through various other CAMs and cytokines. They have a major role in
recognition and binding of immunocompetent cells. This group includes ICAM-1,2 (intercellular adhesion molecule,
also
called CD54), VCAM (vascular cell adhesion molecule, also named CD106), NCAM (neural cell adhesion molecule).
v)CD44. The last group of adhesion molecules is a break away from immunoglobulin superfamily. CD44 molecule binds to
hyaluronic acid and is expressed on leucocytes. It is involved in leucocyte-endothelial interactions as well as in
cell-ECM interactions.
2. CYTOKINES. Another way the cells may communicate with each other is by release of peptides and other
molecules
acting as paracrine function. Cytokines are soluble proteins secreted by haemopoietic and non-haemopoietic cells in
response to various stimuli. Their main role is in activation of immune system. Presently, about 200 cytokines have
been
identified which are grouped in 6 categories:
i) Interferons (IFN)
ii) Interleukins (IL)
iii) Tumour necrosis factor group (TNF, cachectin)
iv) Transforming growth factor (TGF)
v) Colony stimulating factor (CSF)
vi) Growth factors (e.g. platelet-derived growth factor PDGF, epidermal growth factor EGF, fibroblast growth factor
FGF,
endothelial-derived growth factor EDGF, transforming growth factor TGF). Many of these cytokines have further subtypes as alpha, beta, or are identified by numbers. Cytokines involved in leucocyte-endothelial cell interaction are called chemokines while growth factors and other cytokines are named crinopectins.
3. CELL MEMBRANE RECEPTORS. Cell receptors are molecules consisting of proteins, glycoproteins or
lipoproteins and may be located on the outer cell membrane, inside the cell, or may be trans-membranous. These receptor molecules are synthesised by the cell itself depending upon their requirement, and thus there may be upregulation or downregulation of number of receptors. There are 3 main types of receptors:
i)Enzyme-linked receptors. These receptors are involved in control of cell growth e.g. tyrosine kinase associated receptors take part in activation of synthesis and secretion of various hormones.
ii) Ion channels. The activated receptor for ion exchange such as for sodium, potassium and calcium and certain peptide hormones, determines inward or outward movement of these molecules.
iii) G-protein receptors. These are trans-membranous receptors and activate phosphorylating enzymes for
metabolic and synthetic functions of cells.The activation of adenosine monophosphate-phosphatase cycle (c-AMP) by the G-proteins (guanosine nucleotide binding regulatory proteins) is the most important signal system, also known as ‘second messenger’ activation. The activated second messenger (cyclic-AMP) then regulates other intracellular activities. Heat Shock Proteins and Ubiquitin Two proteins which move molecules within the cell cytoplasm are heat shock proteins (HSP) (also called stress proteins) and ubiquitin (so named due to its universal presence in the cells of the body). HSPs. These are a variety of intracellular carrier proteins present in most cells of the body, especially in renal tubular epithelial cells. They normally perform the role of chaperones (house-keeping) i.e. they direct and guide metabolic molecules to the sites of metabolic activity e.g. protein folding, disaggregation of protein-protein complexes and transport of proteins into various intracellular organelles (protein kinesis). However, in response to stresses of various types (e.g. toxins, drugs, poisons, ischaemia), their level goes up both inside the cell as well as they leak out into the plasma, and hence the name stress proteins. In experimental studies HSPs have been shown to limit tissue necrosis in ischaemic reperfusion injury in myocardial infarcts. In addition, they have also been shown to have a central role in protein aggregation in amyloidosis. Ubiquitin. This is another related stress protein which has ubiquitous presence in human body cells. Like HSPs, ubiquitin too directs intracellular molecules for either degradation or for synthesis. Ubiquitin has been found to be involved in a variety of human degenerative diseases, especially in the nervous system in aging e.g. activation of genes for protein synthesis in neurodegenerative diseases such as in Alzheimer’s disease, Creutzfeldt-Jakob disease, Parkinson’s disease.
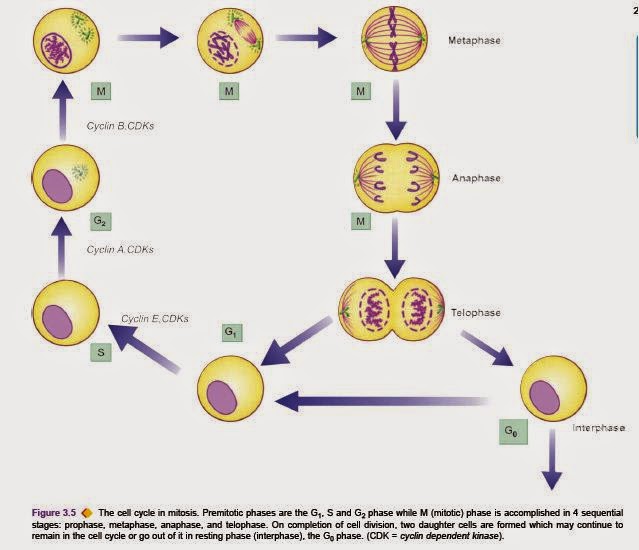
CELL CYCLE
Multiplication of the somatic (mitosis) and germ (meiosis) cells is the most complex of all cell functions. Mitosis is controlled by genes which encode for release of specific proteins molecules that promote or inhibit the process of mitosis at different steps. Mitosis-promoting protein molecules are cyclins A, B and E. These cyclins activate cyclindependent kinases (CDKs) which act in conjunction with cyclins.After the mitosis is complete, cyclins and CDKs are degraded and the residues of used molecules are taken up by cytoplasmic caretaker proteins, ubiquitin, to the peroxisome for further degradation. Period between the mitosis is called interphase. The cell cycle is the phase between two consecutive pisions . There are 4 sequential phases in the cell cycle: G1 (gap 1) phase, S (synthesis) phase, G2 (gap 2) phase, and M (mitotic) phase. G1 (Pre-mitotic gap) phase is the stage when messenger RNAs for the proteins and the proteins themselves required for DNA synthesis (e.g. DNA polymerase) are synthesised. The process is under control of cyclin E and CDKs. S phase involves replication of nuclear DNA. Cyclin A and CDKs control it. G2 (Pre-mitotic gap) phase is the short gap phase in which correctness of DNA synthesised is assessed. This stage is promoted by cyclin B and CDKs. M phase is the stage in which process of mitosis to form two daughter cells is completed. This occurs in 4 sequential stages: prophase, metaphase, anaphase, and telophase (acronym=PMAT).
Prophase. Each chromosome pides into 2 chromatids which are held together by centromere. The centriole pides and the two daughter centrioles move towards opposite poles of the nucleus and the nuclear membrane disintegrates.
Metaphase. The microtubules become arranged between the two centrioles forming spindle, while the chromosomes line up at the equatorial plate of the spindle. Anaphase. The centromeres pide and each set of separated chromosomes moves towards the opposite poles of the spindle. Cell membrane also begins to pide.
Telophase. There is formation of nuclear membrane around each set of chromosomes and reconstitution of the nucleus. The cytoplasm of the two daughter cells completely separates. G0 phase. The daughter cells may continue to remain in the cell cycle and pide further, or may go out of the cell cycle
into resting phase, called G0 phase. Stimulation of mitosis can be studied in a number of ways as under:
Compensatory stimulation of mitosis by removal of part of an organ. Reparative stimulation of mitosis occurs when a tissue is injured.
Target organ stimulation of mitosis occurs under the influence of specific hormones which have mitogenic effect on cells of the target organ.
ETIOLOGY OF CELL INJURY
The cells may be broadly injured by two major ways:
A. By genetic causesB. By acquired causes
. The acquired causes of disease comprise vast majority of common diseases afflicting mankind. Based on underlying agent, the acquired causes of cell injury can be further categorised as under:
1. Hypoxia and ischaemia
2. Physical agents
3. Chemical agents and drugs
4. Microbial agents
5. Immunologic agents
6. Nutritional derangements
7. Aging
8. Psychogenic diseases
9. Iatrogenic factors
10. Idiopathic diseases.
In a given situation, more than one of the above etiologic factors may be involved. These are briefly outlined below.
1. HYPOXIA AND ISCHAEMIA. Cells of different tissues essentially require oxygen to generate energy and perform metabolic functions. Deficiency of oxygen or hypoxia results in failure to carry out these activities by the cells. Hypoxia is the most common cause of cell injury. Hypoxia may result from the following:
The most common mechanism of hypoxic cell injury is by reduced supply of blood to cells due to interruption i.e. ischaemia.However, hypoxia may result from other causes as well e.g. disorders of oxygen-carrying RBCs (e.g. anaemia, carbon monoxide poisoning), heart diseases, lung diseases and increased demand of tissues.
2. PHYSICAL AGENTS.
Physical agents in causation of disease are as under:
- mechanical trauma (e.g. road accidents);
- thermal trauma (e.g. by heat and cold);
- electricity;
- radiation (e.g. ultraviolet and ionising); and
- rapid changes in atmospheric pressure.
3. CHEMICALS AND DRUGS. An ever increasing list of chemical agents and drugs may cause cell injury. Important examples include the following:
- chemical poisons such as cyanide, arsenic, mercury;
- strong acids and alkalis;
- environmental pollutants;
- insecticides and pesticides;
- oxygen at high concentrations;
- hypertonic glucose and salt;
- social agents such as alcohol and narcotic drugs; and
- therapeutic administration of drugs.
4. MICROBIAL AGENTS. Injuries by microbes include infections caused by bacteria, rickettsiae, viruses, fungi, protozoa, metazoa, and other parasites. Diseases caused by biologic agents
5. IMMUNOLOGIC AGENTS. Immunity is a ‘doubleedged sword’—it protects the host against various injurious agents but it may also turn lethal and cause cell injury e.g. hypersensitivity reactions; anaphylactic reactions; and autoimmune diseases. Immunologic tissue injury is discussed in Chapter 4.
6. NUTRITIONAL DERANGEMENTS. A deficiency or an excess of nutrients may result in nutritional imbalances. Nutritional deficiency diseases may be due to overall deficiency of nutrients (e.g. starvation), of protein calorie (e.g. marasmus, kwashiorkor), of minerals (e.g. anaemia), or of trace elements.Nutritional excess is a problem of affluent societies resulting in obesity, atherosclerosis, heart disease and hypertension.
7.AGING. Cellular aging or senescence leads to impaired ability of the cells to undergo replication and repair, and ultimately lead to cell death culminating in death of the inpidual. This aspect is dealt at the end of this chapter.
8. PSYCHOGENIC DISEASES. There are no specific biochemical or morphologic changes in common acquired mental diseases due to mental stress, strain, anxiety, overwork and frustration e.g. depression, schizophrenia. However, problems of drug addiction, alcoholism, and smoking result in various organic diseases such as liver damage, chronic bronchitis, lung cancer, peptic ulcer, hypertension, ischaemic heart disease etc.
9. IATROGENIC CAUSES. Although as per Hippocratic oath, every physician is bound not to do or administer anything that causes harm to the patient, there are some diseases as well as deaths attributed to iatrogenic causes (owing to physician). Examples include occurrence of disease or death due to error in judgment by the physician and untoward effects of administered therapy (drugs, radiation).
10. IDIOPATHIC DISEASES. Idiopathic means “of unknown cause”. Finally, although so much is known about the etiology of diseases, there still remain many diseases for which exact cause is undetermined. For example, most common form of hypertension (90%) is idiopathic (or essential) hypertension. Similarly, exact etiology of many cancers is still incompletely known.
PATHOGENESIS OF CELL INJURY
Injury to the normal cell by one or more of the above listed etiologic agents may result in a state of reversible or irreversible cell injury. The underlying alterations in biochemical systems of cells for reversible and irreversible cell injury by various agents is complex and varied. However, in general, the following principles apply in pathogenesis of most forms of cell injury by various agents:1. Type, duration and severity of injurious agent: The extent of cellular injury depends upon type, duration and severity of the stimulus e.g. small dose of chemical toxin or short duration of ischaemia cause reversible cell injury while large dose of the same chemical agent or persistent ischaemia cause cell death.
2. Type, status and adaptability of target cell: The type of cell as regards its susceptibility to injury, its nutritional and metabolic status, and adaptation of the cell to hostile environment determine the extent of cell injury e.g. skeletal muscle can withstand hypoxic injury for long-time while cardiac muscle suffers irreversible cell injury after 20-30 minutes of persistent ischaemia.
3. Underlying intracellular phenomena: Irrespective of other factors, following essential biochemical Phenomena underlie all forms of cell injury:
i) Mitochondrial damage causing ATP depletion.
ii) Cell membrane damage disturbing the metabolic and trans-membrane exchanges.
iii). Release of toxic free radicals.
4. Morphologic consequences: All forms of biochemical changes underlying cell injury are expressed in terms of morphologic changes. The ultrastructural changes become apparent earlier than the light microscopic alterations. The morphologic changes of reversible cell injury (e.g. hydropic swelling) appear earlier than morphologic alterations in cell death (e.g. in myocardial infarction). The interruption of blood supply (i.e. ischaemia) and impaired oxygen supply to the tissues (i.e. hypoxia) are most common form of cell injury in human beings. genesis of hypoxic and ischaemic cell injury is, therefore, described in detail below followed by brief discussion on pathogenesis of chemical and physical (ionising radiation) agents.